地址:浙江省杭州市西湖区浙江大学紫金港校区
联系人:董老师
邮箱:dong.hy@zju.edu.cn

地址:浙江省杭州市西湖区浙江大学紫金港校区
联系人:董老师
邮箱:dong.hy@zju.edu.cn
神经元的自我回避在无脊椎动物和脊椎动物中都是保守的特征。自我回避的关键机制在于单个神经元中产生一套独特的细胞表面蛋白质,参与同源异构体特异性的嗜同性相互作用,从而驱动神经元的排斥作用而不是粘附作用。在这些细胞表面蛋白中,包括果蝇唐氏综合征细胞粘附分子(Dscam1),脊椎动物成簇的原钙粘蛋白(cPcdhs),以及最近在螯肢动物中发现的截短型Dscam (sDscams)。
近日,我会理事长金勇丰教授团队联合上海交通大学系统生物医学研究院吴强教授团队受邀在Cell Press旗下期刊Trends in Biochemical Sciences上发表题为“Confluence and convergence of Dscam and Pcdh cell-recognition codes”的综述论文,系统性地介绍了节肢动物Dscam(唐氏综合征细胞粘附分子)与脊椎动物cPcdh作为细胞识别码在进化起源、表达调控以及神经元识别中的最新进展。

1. 果蝇Dscam1、脊椎动物Pcdh和螯肢动物sDscam
果蝇Dscam1和脊椎动物的原钙粘蛋白Pcdh是单基因产生极端亚型多样性经典案例,在神经元识别组装中起重要作用。在果蝇中,Dscam1通过互斥可变剪接潜在产生38,016种蛋白异构体。在人类中,串联排列的原钙粘蛋白基因簇(clustered Pcdh)通过可变启动子可以生成53种钙粘蛋白。有趣的是,在螯肢动物中,最近发现了一个“杂合”基因sDscam,该基因在序列上与果蝇Dscam1同源,但在基因组组装上与脊椎动物Pcdh惊人相似。Dscam1和Pcdh能够介导神经元自我与非我识别和自我回避,因此推测螯肢动物sDscam发挥着类似的作用(图1)。
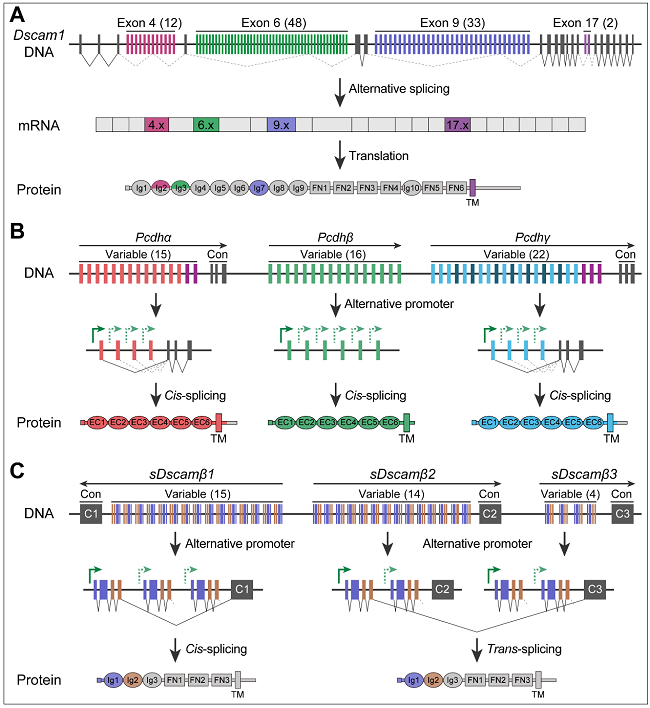
图1. Dscam1、Pcdh和sDscam的基因结构
2. 果蝇Dscam1、脊椎动物Pcdh和螯肢动物sDscam的进化起源
Pcdh和Dscam基因起源于6亿年前左右对称动物和刺胞动物的分化之前,但它们经历了进化枝特异性的亚型多样化扩展。大量多样性的Dscam分子是节肢动物特有的,并且通过不同机制进行多样性扩展。首先,泛甲壳类动物Dscam1基因通过四个可变外显子簇的互斥剪接产生亚型多样性。其次,多足纲物种利用基因复制和互斥剪接来产生Dscam异构体多样性。最后,螯肢动物演化出了大量截短型sDscams。sDscamα和sDscamβ基因的5'端有一系列串联盒式重复序列,分别编码一个和两个Ig结构域,可变盒的数量从50到100不等。在脊椎动物中,不同哺乳动物具有物种特异性扩张和收缩的Pcdh基因。在无脊椎动物章鱼基因组中已鉴定出100多个Pcdhs。相反,Pcdh不存在于蜕皮动物的基因组中,它们进化出了复杂的Dscam来实现其功能,这表明节肢动物和脊椎动物之间存在趋同进化(图2)。
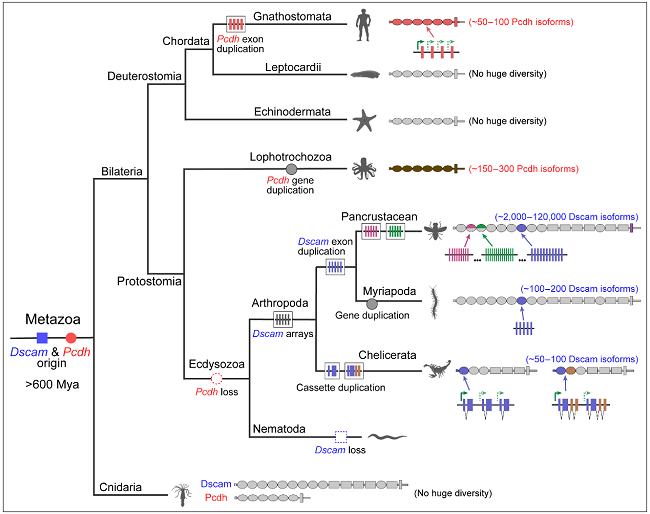
图2. Dscam和Pcdh细胞识别码的进化起源以及多样性扩展
3. 远距离环化介导了Pcdh、Dscam1和sDscam的可变亚型表达
Pcdh的三维基因组结构在其基因表达调控中起重要作用。哺乳动物Pcdh基因簇由Pcdhα和Pcdhβγ两个亚拓扑相关结构域(TAD)组成一个超级TAD。Pcdh的亚TADs由CTCF介导的远端增强子和目标启动子之间的染色质相互作用产生。具体来说,染色质环由ATP驱动的Cohesin蛋白介导的环挤出形成,并优先在正向-反向聚合的CTCF结合位点(CBS)元件之间产生。所有的Pcdh基因簇在远程增强子及其目标启动子中都具有定向CBS元件的串联阵列。
锚定位点和选择序列之间竞争性RNA配对为调控Dscam1的互斥可变剪接提供了一个经典的机制模型。破坏和补偿突变表明,Dscam1可变外显子4、6和9的互斥剪接由锚定位点-选择序列碱基配对调控。最近研究发现sDscamβ的5’可变盒式外显子存在锚定位点和选择序列之间的竞争性RNA配对。破坏和补偿突变表明,单个可变盒式外显子的剪接是由锚定位点-选择序列碱基配对调控的,此外,锚定位点和选择序列分子间的RNA配对调控sDscam的反式剪接(图3)。
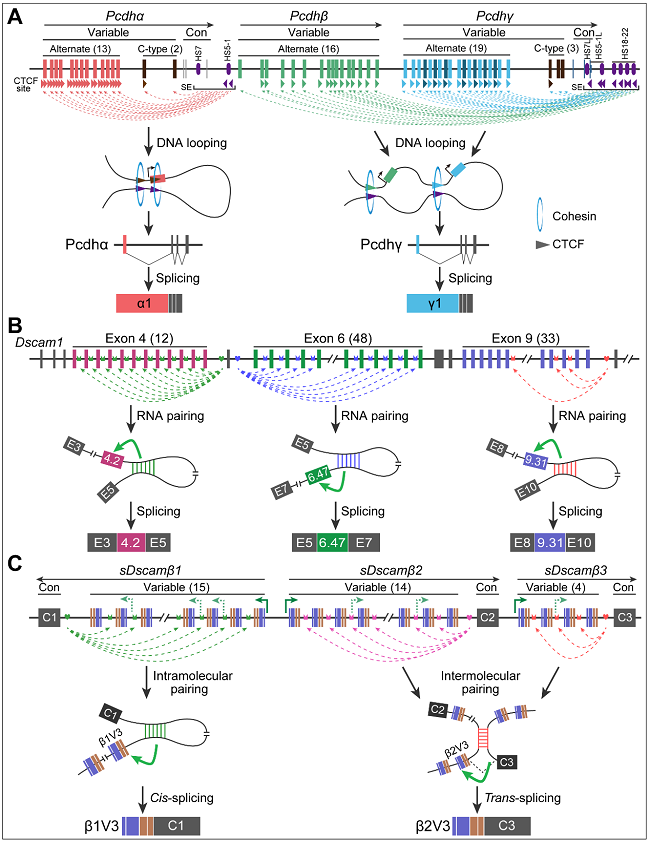
图3. 远距离环化介导了Pcdh、Dscam1和sDscam的表达
4. Dscam1、Pcdh和sDscam表现出严格的同源特异性反式相互作用
果蝇Dscam1、脊椎动物Pcdh和螯肢动物sDscam同种亚型表现出特异性的同源反式结合。在果蝇Dscam1中,这种同源性结合特异性依赖于可变Ig2、Ig3 和 Ig7结构域。可变Ig结构域以模块化方式相互作用,单个可变结构域不匹配也会阻止结合。类似地,Pcdh异构体的顺式二聚体之间的反式相互作用是严格同源的,由胞外域EC1-EC4介导。对于螯肢动物sDscam,同源反式相互作用由可变 Ig1结构域介导,而可变Ig2结构域不会影响同源结合(图4)。
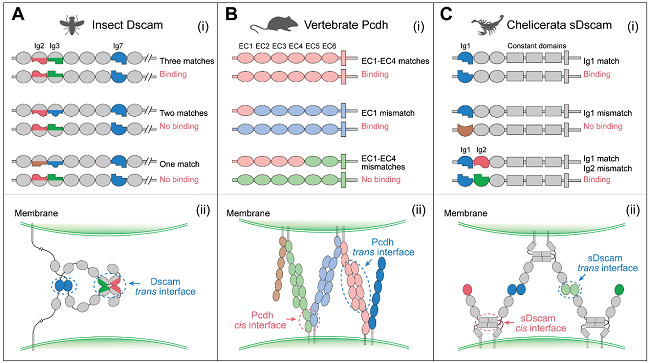
图4. Dscam、Pcdh和sDscam表现出亚型特异性的同源反式结合
5. Dscam1和Pcdh介导神经元自我回避和自我与非我识别
在果蝇中,Dscam1可以产生数万种细胞表面蛋白,每个神经元随机表达不同的Dscam1亚型子集,这些子集表现出高度特异性的同源结合,从而赋予不同神经元不同的细胞识别码。Dscam1缺失导致树突分支神经元广泛的自我交叉,这些缺陷表型可以通过回补任何一种Dscam1亚型来挽救。然而,将Dscam1的多样性减少到396或576种亚型时,I类和III类树突分枝神经元之间的树突重叠减少。因此,虽然表达单一Dscam1亚型足以维持树突分支神经元的自我回避,但需要数千种Dscam1亚型来区分自我和非我神经元。与果蝇Dscam1类似,脊椎动物Pcdhs的条件性敲除会导致星形胶质细胞的树突塌陷,这是一种典型的自我回避缺陷。此外,三个Pcdh簇的缺失会导致小鼠嗅觉感觉神经元严重的轴突分枝缺陷和自我回避的丧失。此外,与仅删除Pcdhγ相比,Pcdhα/γ簇的缺失导致更严重的生存和自我回避缺陷。因此,Pcdh作为细胞表面识别码,可以介导脊椎动物的自我回避和自我与非我识别。最近在皮层神经元中的研究报道了Pcdh介导同源结合的潜在胞内信号通路(图5)。
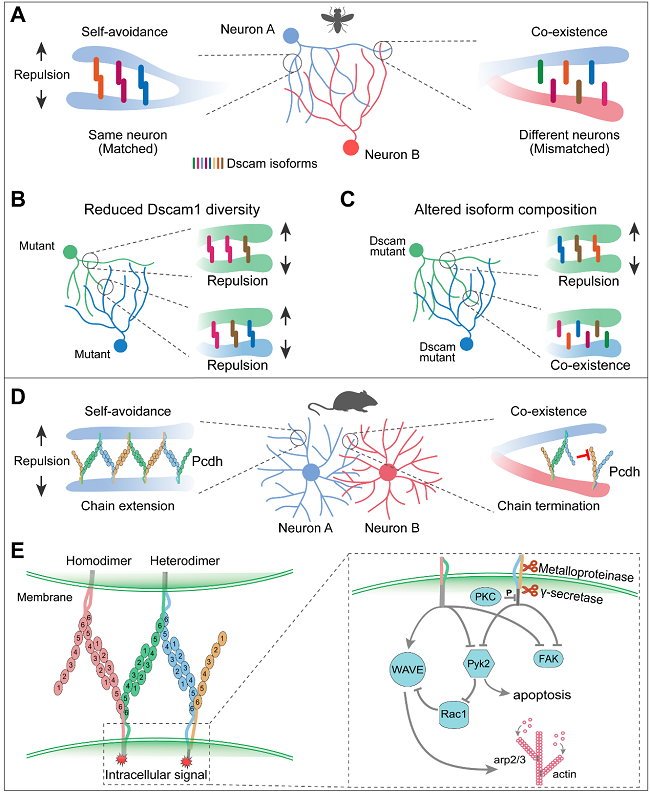
图5. Dscam1和Pcdh介导自我回避和自我与非我识别
6. 非自我回避功能:挑战传统观点
多种遗传证据证实了Dscam1介导树突分枝神经元的自我回避模型,但自我回避是否是Dscam1在蘑菇体(MB)轴突分叉中的功能机制仍存在争议。在传统模型中,MB神经元的姐妹分支轴突分离依赖于Dscam1介导的排斥信号。然而,最近的遗传学证据发现了Dscam1突变体表现出新的缺陷表型:MB叶截短和两叶变细,这些表型缺陷不能用Dscam1介导的自我回避解释。此外,最近的Dscam1表型-多样性相关性分析显示,Dscam1多样性对树突分枝神经元、蘑菇体和机械感觉神经元发育模式的需求不同。这些结果表明Dscam1通过不同的机制在轴突和树突中起着不同的作用。除了自我回避和自我与非我识别功能外,Pcdhα簇的成员还调节皮层神经元的迁移。此外,Pcdhγ簇的缺失会导致小鼠脊髓中间神经元死亡和新生小鼠致死。Pcdhγ促进触摸感觉神经元的外周轴突分支。因此,果蝇Dscam和哺乳动物Pcdh在神经元发育中也表现出非自我回避功能(图6)。
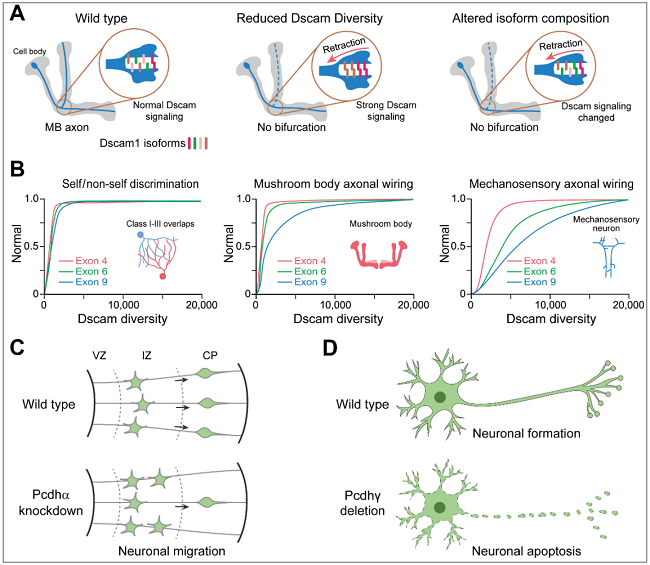
图6. 果蝇Dscam和哺乳动物Pcdh的非自我回避功能
总结与展望
本文中,作者首先介绍了Dscam和Pcdh在节肢动物和脊椎动物之间存在趋同进化并阐明了其可变启动子选择以及顺式、反式可变剪接的调控机制。然后,介绍了Pcdh和Dscam介导嗜同性相互作用的分子结构基础,回顾了其作为细胞识别码调节神经元自我与非我识别的作用模式。最后,总结了最近发现的Pcdh和Dscam在发育过程中表现出的非自我回避功能。然而,仍存在一些问题有待进一步研究,如:Dscam/sDscam和Pcdh如何在单个细胞中进行随机表达和动态调控仍不清楚。Dscam和Pcdh介导的同源反式相互作用如何转化为接触依赖的神经突排斥信号还有待进一步阐明。果蝇Dscam1、螯肢动物sDscam和脊椎动物Pcdh基因在其他非神经元组织中表达,可能还具有其他非神经元功能。
金理事长和上海交通大学吴强教授为论文的通讯作者,金勇丰教授课题组博士后董海洋和上海交通大学李金环博士为论文的共同第一作者。该论文获得了国家自然科学基金、国家重点研发计划项目、浙江省自然科学基金、浙江大学上海高等研究院繁星科学基金和上海市科学技术委员会资助。
原文链接:https://www.cell.com/trends/biochemical-sciences/fulltext/S0968-0004(23)00227-X





